Structural and functional brain differences between dyslexic and typical readers are discussed. What do these differences mean for people with dyslexia?
Table of contents:
- Introduction
- Neuroplasticity: An extraordinary discovery
- Brain differences in the dyslexia brain
- Poor connectivity in the dyslexia brain
- Do brain differences equal brain disorders?
- Conclusion
- Key takeaways
Introduction

Developmental dyslexia is considered a neurodevelopmental learning disability, estimated to affect between 5 and 13% of the U.S. population. The hallmark behavioral profile of dyslexia is difficulty with decoding and recognition of words that cannot be accounted for by classroom instruction, motivation, or overall cognitive abilities.
Ideas connecting dyslexia to brain functioning or brain lesions were already conveyed in the late 19th century by Berlin (Opp, 1994), Morgan (1896), and Hinshelwood (1895). Autopsy studies in the 1980s and 90s of persons with documented histories of dyslexia seemingly confirmed these long-held beliefs.
Specifically, Galaburda and colleagues (1985) reported several anatomical anomalies in the brains of a few persons with dyslexia. Although those findings are intriguing, they are far from decisive. The samples were extremely small (a total of four men and three women) and included participants with evidence of neurological or psychiatric conditions and with impairments not limited to written language, who should have been excluded from a study purporting to examine brain correlates of dyslexia. The control group was also very small, effectively precluding reliable estimation of the specificity of any findings.
However, people were also convinced that the brain was hardwired at the time and during the preceding decades. The prevailing theory was that nothing could be done if part of the brain was damaged due to injury, or if a person was born with a mental condition (Keshav, 2018). This certainly contributed to the opinion that dyslexia is a lifelong condition — “like alcoholism … it can never be cured” (Clark & Gosnell, 1982).
Neuroplasticity: An extraordinary discovery
A 1998 landmark study found that the human brain can generate new neurons (Eriksson et al., 1998). This research challenged the prevailing theory that the human brain is a rigid system, that humans are born with all their brain cells, and that we lose brain cells daily, which our brain does not replace with new ones. More research studies followed, demonstrating that the brain is malleable and adaptable — or plastic — and even that it can grow.
In a classic experiment, Maguire et al. (2000) compared the brains of London taxi drivers with a control group of non-taxi drivers. Maguire first got the idea to study London cab drivers from research on the memory champions of the animal world. Some birds and mammals, such as western scrub jays and squirrels, cache food and dig it up later, which means they must memorize the locations of all their hiding spots.
Researchers noticed that the hippocampus, which is crucial for long-term memory and spatial navigation, was much larger in these animals than in similar species that did not secret away their snacks. In some species, hippocampal volumes enlarge specifically during seasons when the demand for spatial ability is greatest. This interesting fact led Maguire to wonder whether the hippocampus would grow in people who had to memorize many visual locations, such as London cab drivers who have to memorize roughly 25,000 city streets, as well as thousands of tourist attractions and hot spots, before earning their cab licenses.

The results showed that London taxi drivers had significantly larger posterior hippocampi than the controls (Maguire et al., 2000) and London bus drivers (Maguire et al., 2006). Contrary to taxi drivers, who must navigate through various routes around the city, bus drivers have to follow only a limited, predetermined set of routes. Furthermore, years of navigation experience correlated with hippocampal gray matter volume in taxi drivers.
Similar findings of apparently environmentally driven plasticity, with positive correlations between gray matter and the time spent learning and practicing their specialization, have been reported in several other groups, including musicians (Sluming et al., 2002), jugglers (Draganski et al., 2004), and bilinguals (Mechelli et al., 2004).
The studies mentioned above and many others confirm that the brain can change and even that an injured brain can be rewired. The healthy portions of a damaged brain can be trained to assume the functions of the dysfunctional tissue (Nudo, 2013; Kadosh & Walsh, 2006).
Brain differences in the dyslexia brain
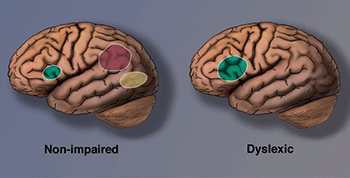
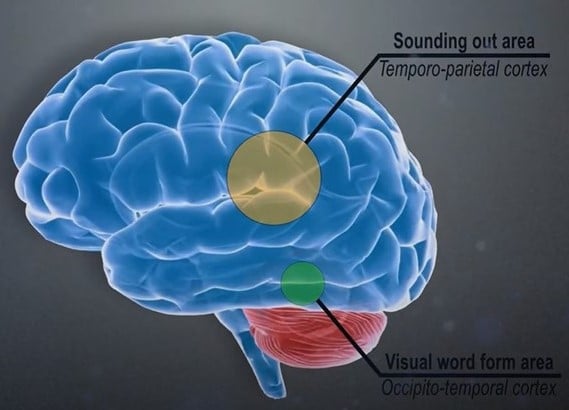
As technology advanced, neuroscience contributed more and more to dyslexia research. Unfortunately, most studies have too small samples to permit reliable conclusions, and many results are inconsistent (Protopapas & Parrila, 2018). In a meta-analysis of functional neuroimaging studies of dyslexia, Martin et al. (2016) list studies in which differences between groups with and without dyslexia were found in specific brain regions. The most consistent findings concerned the left occipitotemporal cortex, which includes the so-called visual word form area (VWFA), thought critical for reading.
Neuroscientists at Georgetown University Medical Center concluded that skilled readers can recognize words at a lightning-fast speed when they read because the words have been placed in a sort of visual dictionary. This part of the brain, the VWFA (shown in yellow), functions separately from an area that processes the sounds of written words (Glezer et al., 2016).
Glezer and her coauthors tested word recognition in 27 volunteers in two different experiments using fMRI. The researchers could see that words that are different but sound the same — like ‘hare’ and ‘hair’ — activate different neurons, akin to accessing different entries in a dictionary’s catalog. If the sounds of the word had any influence on this part of the brain, we would expect to see that they activate the same or similar neurons. This, however, was not the case — ‘hair’ and ‘hare’ looked just as different as ‘hair’ and ‘soup.’
In addition, the researchers found a distinct region sensitive to sounds, where ‘hair’ and ‘hare’ did look the same. The researchers thus showed that the brain has regions that specialize in doing each of the components of reading: one region is doing the visual piece, and the other is doing the sound piece.
In the largest study of its kind up to date, Brem et al. (2020) confirmed that the VWFA is key to fluent reading using fMRI data (n = 140 children, 7.9–12.2 years; 55 children with dyslexia, 73 typical readers, 12 intermediate readers). Their study also showed that connectivity of the left occipitotemporal cortex seems a prerequisite for guiding the emerging specialization in the VWFA. Furthermore, Pedago et al. (2011) supported the VWFA’s crucial role in the discrimination of letter orientation, which links to the well-known fact that a deficit in discriminating between a b and a d is a common symptom of dyslexia.
Given that the most robust underactivation in children with dyslexia occurs in the left occipitotemporal cortex (Brem et al., 2020; Martin et al., 2016), this brain region should be a prime target for supportive interventions.
These findings negate the dual-access theory, which suggests that visual word recognition in skilled readers is not based on visual processing alone but that we access both the phonology and the visual perception of a word (Frost, 1998).
The left inferior parietal lobule came in a close second in the meta-analysis study by Martin and colleagues (shown in red). This part of the brain is said to be involved in word analysis, grapheme-to-phoneme conversion, and general phonological and semantic processing.
Imaging also reveals compensatory overactivation in other parts of the reading system (shown in green). The compensatory neural systems allow a dyslexic person to read more accurately. However, the critical visual word-form area remains disrupted, and difficulties with rapid, fluent, automatic reading persist. The dyslexic continues to read slowly (Shaywitz, 2005).
Poor connectivity in the dyslexia brain
According to a team of scientists at the MRC Cognition and Brain Sciences Unit, University of Cambridge, there are, in fact, no specific brain areas that cause learning disabilities, such as dyslexia (Siugzdaite et al., 2020).
To test their hypothesis, the researchers used machine learning to map the brain differences across a group of almost 479 children, 337 of whom had been referred with learning-related cognitive problems and 142 from a comparison sample. The algorithm interpreted data from a large battery of cognitive, learning, and behavioral measures, and from brain scans taken using magnetic resonance imaging (MRI).
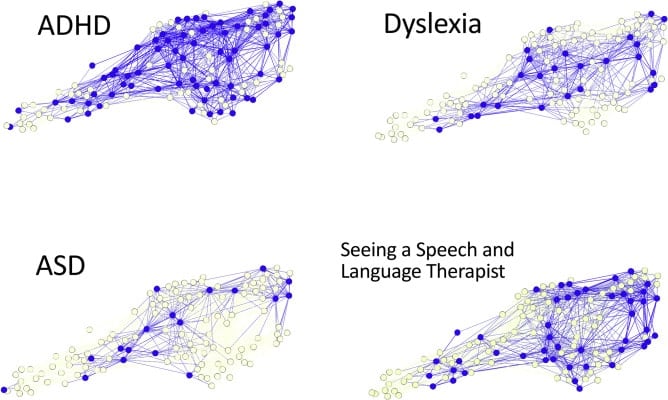
The researchers found that the brain differences did not map onto any labels the children had been given — in other words, there were no brain regions that predicted having ADHD, dyslexia, or ASD (autism spectrum disorder). More surprisingly, they found that the different brain regions did not even predict specific cognitive difficulties — there was no specific brain deficit for language or memory difficulties, for example.
Instead, the team found that the children’s brains were organized around hubs, like an efficient traffic system or social network. Children with well-connected brain hubs had very specific cognitive difficulties, such as poor listening skills, or no cognitive difficulties. By contrast, children with poorly connected hubs — like a train station with few or poor connections — had widespread and severe cognitive problems.
Do brain differences equal brain disorders?
While Protopapas and Parrila (2019) stress that poor reading is a serious problem that requires special attention in the form of assessment and remediation as early as possible, they point out that brain differences do not equal brain disorders. After reviewing the scientific evidence of the last few decades, Protopapas and Parrila (2018) conclude:
Differences in brains are certain to exist whenever differences in behavior exist, including differences in ability and performance. Therefore, findings of brain differences do not constitute evidence for abnormality; rather, they simply document the neural substrate of the behavioral differences. We suggest that dyslexia is best viewed as one of many expressions of ordinary ubiquitous individual differences in normal developmental outcomes. Thus, terms such as “dysfunctional” or “abnormal” are not justified when referring to the brains of persons with dyslexia.
It should also be noted that brain differences may be the result of reading difficulties and not the cause. In one study, published online in the Journal of Neuroscience, researchers analyzed the brains of children with dyslexia. They compared them to two other groups of children: an age-matched group without dyslexia and a group of younger children who had the same reading level as the children with dyslexia.
Although the children with dyslexia had less gray matter than age-matched children without dyslexia, they had the same amount of gray matter as the younger children at the same reading level. Krafnick et al. (2014) say this suggests that the anatomical differences reported in left-hemisphere language-processing regions of the brain appear to be a consequence of reading experience as opposed to a cause of dyslexia.
Huettig et al. (2018) highlight that most known deficits associated with reading impairments also occur in “normal” illiterate or low-literate adults who have received no reading instruction or very little. They argue that in a substantial number of individuals with dyslexia, many deficiencies are secondary consequences of a lack of (adequate) reading experience. They refer to a study by Skeide et al. (2016) that discovered that when adults learn to read for the first time, the changes that occur in their brain are not limited to the outer layer of the brain, the cortex, but extend to deep brain structures in the thalamus and the brainstem. This study observed illiterate Indian women who learned how to read and write for six months.
Conclusion
There are indeed brain differences between people with dyslexia and typical readers. If these brain differences are the cause or the result of dyslexia is not conclusive. We know, however, that the brain is plastic. In fact, the human brain is a powerhouse; the human brain has put a man on the moon, created the silicon chip that can do billions of calculations per second, invented red, yellow, and green lights to control millions of people in traffic every day and — believe it or not — found ways to see what goes on inside itself. The human brain itself tells us that it is most certainly capable of overcoming learning obstacles like dyslexia, despite brain differences.
We know that at least two brain areas are involved in learning to read: the visual word form area (VWFA) and the left parietal lobe. Both must be trained in the teaching of reading and spelling, and Edublox’s Live Tutor aims to do just that. Our program is based on the Orton-Gillingham approach, which is excellent at developing the left parietal lobe (the sound piece), but our program simultaneously targets the brain’s VWFA. The VWFA remains highly malleable into adulthood (Martin et al., 2019).
Practicing “nonsense words” does not feature in Edublox programs as this counters developing the VWFA. “Nonsense words” are letter sequences that follow regular phonetic rules and are pronounceable but have no meaning — for example, bif, yom or mig. Many schools have implemented tools to measure early reading ability, which include tests of the ability to decode nonsense words (Marshall, 2019). However, it has now also become a skill that is increasingly being taught directly, especially in Orton-Gillingham programs.
We also know that connectivity, especially in the VWFA, is a prerequisite to learning to read. Edublox’s Development Tutor works in conjunction with Live Tutor and aims to improve cognitive skills — and, ultimately, brain connectivity.
Watch our playlist of customer reviews and experience how Edublox training and tutoring help turn dyslexia around.
Key takeaways
Authored by Sue du Plessis (B.A. Hons Psychology; B.D.), an educational specialist with 30+ years’ experience in the field of learning disabilities.
Medically reviewed by Dr. Zelda Strydom (MBChB).
References:
Brem, S., Maurer, U., Kronbichler, M. et al. (2020). Visual word form processing deficits driven by severity of reading impairments in children with developmental dyslexia. Scientific Reports, 10.
Clark, M., & Gosnell, M. (1982, March 22). Dealing with dyslexia. Newsweek: 55-6.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., & May, A. (2004). Neuroplasticity: Changes in grey matter induced by training. Nature, 427: 311-2.
Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A., Nordborg, C., Peterson, D. A., Gage, F. H. (1998). Neurogenesis in the adult human hippocampus. Natural Medicine, 4(11): 1313-7.
Frost, R. (1998). Toward a strong phonological theory of visual word recognition: True issues and false trails. Psychological Bulletin, 123(1): 71-99.
Galaburda, A. M., Sherman, G. F., Rosen, G. D., Aboitiz, F., & Geschwind N. (1985). Developmental dyslexia: Four consecutive patients with cortical anomalies. Annals of Neurololgy, 18(2): 222-33.
Glezer, L. S., Eden, G., Jiang, X., Luetje, M., Napoliello, E., Kim, J., & Riesenhuber, M. (2016). Uncovering phonological and orthographic selectivity across the reading network using fMRI-RA. Neuroimage, 138: 248-56.
Hinshelwood, J. (1895). Word-blindness and visual memory. Lancet, 146(3773): 1564-70.
Huettig, F., Lachmann, T., Reis, A., & Petersson, K. M. (2018). Distinguishing cause from effect—Many deficits associated with developmental dyslexia may be a consequence of reduced and suboptimal reading experience. Language, Cognition and Neuroscience, 33(3): 333–50.
Kadosh, R. C., & Walsh, V. (2006). Cognitive neuroscience: Rewired or crosswired brains? Current Biology, 16(22): R962-3.
Keshav, M. (2018). The life transforming power of NLP. Chennai: Notion Press.
Krafnick, A. J., Flowers, D. L., Luetje, M. M., Napoliello, E. M., & Eden, G. F. (2014). An investigation into the origin of anatomical differences in dyslexia. Journal of Neuroscience, 34(3).
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S. J., & Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences of the United States of America, 97(8): 4398-403.
Maguire, E. A., Woollett, K., & Spiers, H. J. (2006). London taxi drivers and bus drivers: A structural MRI and neuropsychological analysis London taxi drivers and bus drivers. Hippocampus, 16(12): 1091-101.
Marshall, A. (2023). The nonsense of teaching nonsense words. Dyslexia.com
Martin, A., Kronbichler, M., & Richlan, F. (2016). Dyslexic brain activation abnormalities in deep and shallow orthographies: A meta-analysis of 28 functional neuroimaging studies. Human Brain Mapping, 37: 2676–99.
Martin, L., Durisko, C., Moore, M. W., Coutanche, M. N., Chen, D., & Fiez, J. A. (2019). The VWFA is the home of orthographic learning when houses are used as letters. eNeuro, 6(1).
Mechelli, A., Crinion, J. T., Noppeney, U., O’Doherty, J., Asburner, J., Frackowiak, R. S., & Price, C. J. (2004). Structural plasticity in the bilingual brain. Nature, 431: 757.
Morgan, W. P. (1896). A case of congenital word blindness. The Lancet, 2(1871): 1378.
Nudo, R. J. (2013). Recovery after brain injury: Mechanisms and principles. Frontiers in Human Neuroscience, 7(887).
Opp, G. (1994). Historical roots of the field of learning disabilities: Historical roots of the field of learning disabilities: Some nineteenth-century German contributions. Journal of Learning Disabilities, 27: 10-9.
Pedago, F., Nakamura, K., Cohen, L., & Dehaene, S. (2011). Breaking the symmetry: Mirror discrimination for single letters but not for pictures in the Visual Word Form Area. Neuroimage, 55: 742-9.
Protopapas, A., & Parrila, R. (2018). Is dyslexia a brain disorder? Brain Sciences 8(4).
Protopapas, A., & Parrila, R. (2019). Dyslexia: Still not a neurodevelopmental disorder. Brain Sciences, 9(1).
Shaywitz, S. (2005). Overcoming dyslexia. New York: Vintage Books.
Siugzdaite, R., Bathelt, J., Holmes, J., & Astle, D. E. (2020). Transdiagnostic brain mapping in developmental disorders. Current Biology, 30(7).
Skeide, M. A., Kumar, M., Mishra, R. K., Tripathi, V. N., Guleria, A., Singh, J.P., … & Huettig, F. (2017). Learning to read alters cortico-subcortical cross-talk in the visual system of illiterates. Science Advances, 3(5).
Sluming, V., Barrick, T., Howard, M., Cezayirli, E., Mayes, A., & Roberts, N. (2002). Voxel-based morphometry reveals increased gray matter density in Broca’s area in male symphony orchestra musicians. Neuroimage, 17(3): 1613–22.

